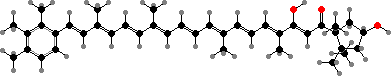
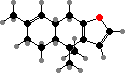
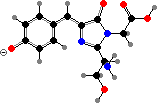
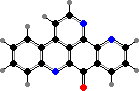
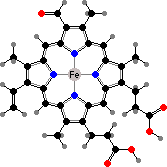
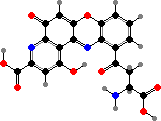
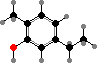
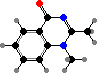
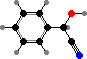
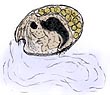
Trikentriorhodin
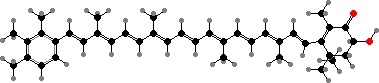
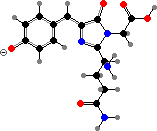
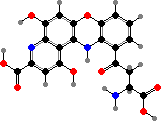
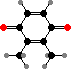
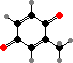
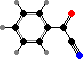
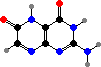
Clathriaxanthin
Introduction -
Common -
Bacteria -
Plantae -
Chromista -
Protozoa -
Fungi -
Animalia -
References
Porifera -
Cnidaria -
Echinodermata -
Chordata -
Annelida -
Mollusca -
Arthropoda
Animals are nearly all multi-cellular, though varying from less than a millimetre to dozens of metres in length. The cells are held together by a protein matrix without rigid walls, and are generally organized around a central chamber or tube, which in sponges is the exit for a canal system and in others serves to hold and digest food.
Sponges are mainly marine and sedentary, often forming colonies of indefinite shape and with cells generally capable of changing form and position. Food is strained out from water taken in through a system of pores and canals, driven by flagellate cells with collars like in choanoflagellate protozoa. Most also have a supportive skeleton, commonly with mineral spicules.
The largest group are the demosponges. In these the skeleton is often a network of collagen fibres called spongin, variously without, with, or replaced by spicules. The latter are formed from silica around one or four axes and include both megascleres and smaller microscleres. They usually produce solid parenchymella larvae, though there are also hollow sorts.
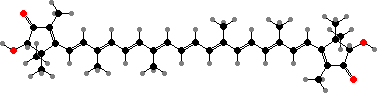
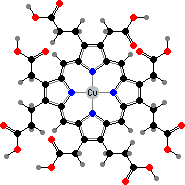
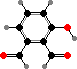
Many demosponges have yellow to purple colours from carotenoids, mainly β-carotene, zeaxanthin, and kinds with aromatic rings like trikentriorhodin and isorenieratene. The last is also found in bacteria, while others appear related to carotenoids from algae. Clathriaxanthin, the main pigment in some Poecilosclerida like red Clathria, is likewise related to astaxanthin.
Several alkaloids are scattered among unrelated genera and sometimes other animals. The trisoxazoles include halichondramide from Chondrosia, Jaspis, and Halichondria, and mycalolides from Mycale, all in different orders; mycalolides are also found in the stony corals Tubastrea. These bind to microfilaments and deter various predators and fungi.
Another sort are pyridoacridines, which are mostly cytotoxic and vary in colour with pH. Here kuanoniamines occur in both Stelletta and Oceanapia along with related compounds. These have been found localized in sponge cells rather than bacteria, but even so they also occur among sea squirts and their predators.
Verongiida have collagenous skeletons lacking spicules except for Hexadella with neither. The spongin fibres have a characteristic pith of dark cellular material and defined outer bark, and the larvae are hollow and develop in external eggs. Many kinds are aerophobic, becoming dark purple or black when exposed to air.

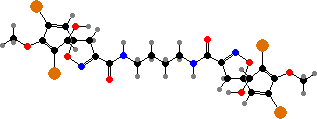
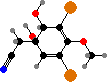
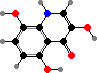
Bromotyrosine derivatives are also typical of this group. Aplysina for instance have isoxazolines like aerothionin, partly an activated defense forming aeroplysinin-1 on injury, but also themselves deterring various bacteria, predators, and settlement. There are also some quinolines like yellow uranidine, which is what reacts with oxygen in A. aerophoba.
Dictyoceratida mostly have skeletons of spongin in dense networks, commonly with two or three sizes of fibres, including the traditional bath sponges. The few exceptions are Vaceletia and fossil relatives which instead form series of calcified chambers. Sponges here typically have parenchymellae larvae and lack cellular pith or basal plates.
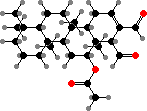
Terpenoids are common in this group and the related Dendroceratida. Some representative types here are scalaradial in Cacospongia, concentrated toward tips, and furodysinin along with similar kinds in dysideids. These deter predators like fish, and sometimes also microbes or competing sponges.
Poecilosclerida are a diverse group with both spicules and spongin showing regional differentiation. The former take the shape of single-axis megascleres and various straight or curved microscleres, which in most kinds include chelae, a unique bent form that have inward spines at the ends. They also share parenchymella larvae with uniform cilia.
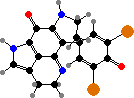
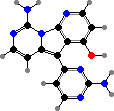
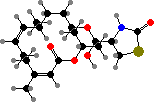

A variety of cytotoxic alkaloids are found here. Several kinds have pyrroloquinolines like green discorhabdins in Latrunculia, or guanidines like purple variolins in Kirkpatrickia, both of which are antimicrobial and deter predatory starfish. Another notable sort are latrunculins in Negombata magnifica, which render them extremely toxic to fish.
Cnidarians have bodies with a jelly-like middle layer and no central nervous, circulatory, or excretory systems. They typically have radial symmetry around the mouth, which is the single opening to the digestive chamber, surrounded by tentacles with stinging cells. Different stages may take the form of tube-like polyps, free-swimming medusae, or more complex colonies.
A variety of cnidarians are bioluminescent, probably mostly to warn off predators though sometimes to lure prey. Like many other animals this comes from oxidizing coelenterazine obtained in their diet. This emits blue light in scyphozoans but in others excites green fluorescent proteins or their variants, changing the colour and sometimes increasing the yield.

These proteins are special in forming their own chromophores, a tyrosine and adjacent residues oxidizing into an imidazolinone ring; GFP in Aequorea and ptilGFP in Ptilosarcus are representative. They are also found in non-luminescent anthozoans, as well as some copepods and lancelets, and since they come from single genes have been useful in fields like live cell imaging.
Anthozoa are exclusively marine polyps but with relatively complex structure. The stomach is divided by radial walls extending to a tubular pharynx, which typically has one or more ciliated channels that take in water. Some are solitary but many form colonies, varying from slow crawling or burrowing types to ones with rigid skeletons.
Soft and horny corals contain a variety of defensive terpenoids. Among these, blue guaiazulene and derivatives give colour to some species like Euplexaura erecta, though less common pigments than reddish polyenes or astaxanthin. Pseudopterosins from Antillogorgia elisabethae are notable for use as an anti-inflammatory.
Astaxanthin or derivatives are also common as fatty acid esters in sea anemones, notably purple-red actinioerythrol, which gives colour to various types like Actinia and Urticina. The red or purplish marks in Calliactis parasitica, found on shells with hermit crabs, are on the other hand from the unique pyridoacridine pigment calliactine.
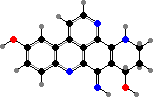
Many anthozoans have not just green fluorescent proteins but variants, with emission shifted to cyan or yellow or with modified chromophores. The most conspicuous in white light are blue to pink chromoproteins, also called pocilloporins, found for instance in sea anemone tentacles and growing tips or coloured morphs of stony corals.
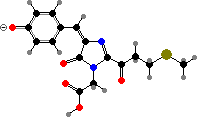
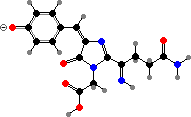
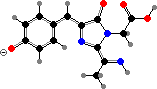
These proteins mature by further oxidation, breaking the peptide chain in pinkish asulCP from Anemonia and forming an imine link in others. The chromophores end up twisted and so non-fluorescent; related isomers with a planar configuration give red fluorescent proteins. These two types are for instance represented in meffCP and meffRFP from Montipora.
The remaining animals all have more complex structures derived from three embryonic layers, of which the middle often gives rise to an internal space or coelom. For the most part, they have bilateral symmetry, which typically goes with a complete digestive tract and often some degree of cephalization. There are exceptions, though, of which the echinoderms are the most notable.
Echinoderm larvae have bilateral symmetry, but the adults instead typically have 5-fold radial symmetry about the mouth. Nearly all are marine and benthic. They have a developed coelom and characteristic water vascular system, which typically supports a series of muscular tube feet used in movement and feeding, as well as an internal calcareous skeleton.
Crinoidea have a cup-shaped body with the mouth surrounded by feather-like arms, often divided into ten to hundreds of branches, each lined with two rows of short pinnules. These are held upward to capture food particles, carried in by mucus in grooves. Most orders have stalks but they are lost in feather-stars.
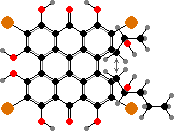
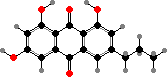
Quinones are common here. Several stalked crinoids are coloured green by gymnochromes, and this same ring structure is preserved in purple fringelites found in some Jurassic fossils. Feather-stars mainly have anthraquinones like crinemodin and derivatives giving red to purple colours. Both often occur in the form of sulfates shown to repel fish.
Echinoidea, the sea urchins, are globular or disc-shaped with movable spines and tube feet extending over most of the body. The skeleton is fused together into a thin test. They move with the mouth on the bottom, usually with five protrusible teeth that scrape up food; most are herbivores but there are also some scavengers or detritus feeders.
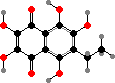
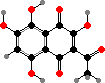
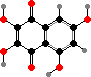
The main pigments here are naphthoquinones, also known in a few other echinoderms. Red echinochrome A occurs in various tissues but others are restricted to the spines and test, where they take the form of calcium salts. The most common are spinochromes A and B, associated with purple and green colours, and their hydroxy derivatives.
Holothuroidea, the sea cucumbers, have elongate bodies with the skeleton mostly reduced to tiny ossicles. The tube feet occur in rows along its length or are sometimes absent, and there is also a ring of 8 or more feeding tentacles around the mouth, which variously trap suspended particles or pick up detritus.
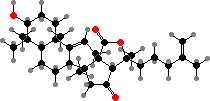
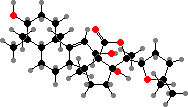
Saponins are major defensive toxins in sea cucumbers and starfish. Here these are glycosides of C30 terpenoids, for instance holotoxigenol from various stichopodids, psolids, and cucumariids. In many holothuriids, where holothurigenol is common, saponins are concentrated in Cuvierian tubules expelled from the anus as sticky strings to repel predators.
Asteroidea and Ophiuroidea, the starfish and brittle stars, have bodies with five or more arms radiating out from a central disc with the mouth underneath. Starfish are mainly carnivores with eversible stomachs and tube feet in open grooves, while brittle stars have articulated arms and are most often scavengers or detrivores.
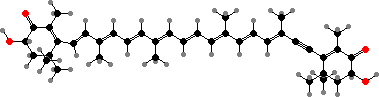
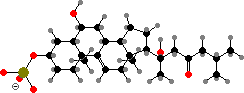
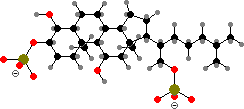
Many starfish are brightly coloured by carotenoids like astaxanthin or didehydroastaxanthin, often in the form of red to blue protein complexes. They deter predators with steroid saponins, of which thornasterol A sulfate is the most common. Brittle stars also produce deterrent steroids but mostly polysulfates like the cholestanetetrol derivative shown.
Chordates have at least an embryonic or larval stage with a stiffened notochord, which runs along the body into a tail, though it is commonly lost in adults. Most also share a dorsal nerve cord and pharyngeal arches, which are variously developed into gills or structures for feeding.
Tunicata are marine, including both plankton and the sessile sea squirts. Here the pharynx takes the form of a mucus net that captures food particles. A few retain a tail and notochord but most lose them to become sac-like adults, which filter water driven between two siphons by cilia. They are hermaphrodites but some also form budding colonies.
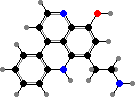
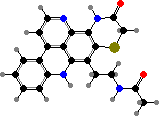
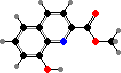
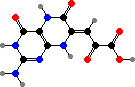
Pyridoacridines are common in a few families and give some red to blue colours. Among styelids there are styelsamines in Eusynstyela. The polycitorids Cystodytes have various types like ascididemin or purple shermilamines and kuanoniamines, and the former two are also representative of didemnids, respectively in Didemnum and Trididemnum.
Actinopterygii, the ray-finned fish, and the following classes are all vertebrates – the notochord is generally replaced by a vertebral column forming part of an internal skeleton. In most this also includes an arch-derived jaw and two sets of paired fins or limbs. Many kinds breathe with paired lungs in addition to gills, or more often instead of them.
Ray-finned fish themselves have gills and are common in most aquatic habitats. The name refers to the fins mostly having no fleshy lobes, so consisting of bony rays supporting a web of skin. Bichirs have simple lungs but in others they are replaced by a more dorsal gas bladder, generally involved in maintaining buoyancy and only occasionally respiration.
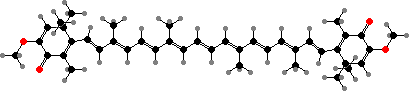
Carotenoids are again common pigments, with mixed epimers found in ester form. Astaxanthins are typical reds, which stand out in many colourful freshwater fish, but in the ocean mostly act as dark shading since longer wavelengths are quickly absorbed. Bright markings are then more often from yellow carotenoids like tunaxanthins.
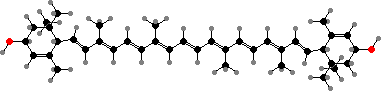
Many marine fish also fluoresce in blue light, varying from camouflage against objects like corals to a visual signal. This was reported in Kaupichthys as from protein-bound bilirubin, a derivative of waste biliverdin in various vertebrates. Biliverdin itself sometimes colours tissues like blood in wrasses and likely blue-green epidermis in Etheostoma darters.
Amphibia as adults mostly breathe air through lungs or their skin, which is permeable with mucous and granular glands, but the young usually take the form of freshwater larvae with external gills. Living groups also share some skeletal reductions like partial ribs and front limbs with at most four digits. Nearly all are predators and depend on moisture for their eggs.
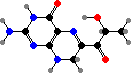
Many salamanders and frogs like Ambystoma, Plethodon, and Lithobates have bright colours from pterins rather than carotenoids. The most prominent are yellow sepiapterin and red drosopterin enantiomers. The latter are also the main red pigments in many reptiles like anoles and snakes, plus a few fish like poeciliids and the eyes of small fruit flies.
Here such colours often serve to advertize skin toxins, which are found in the granular gland secretions from a variety of amphibians. Among Salamandridae many freshwater newts have tetrodotoxin, as do a few frogs. The terrestrial Salamandra (fire salamanders) on the other hand produce samandarine, a neurotoxin causing convulsions.
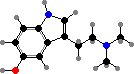

Many frog families including toads and tree frogs secrete serotonin and analogues like bufotenine, mainly as irritants, with effects like vasoconstriction and some psychoactivity. Some also have peptide toxins. Dendrobatidae (poison dart frogs) accumulate different alkaloids from prey like insects or mites, notably pumiliotoxins, histrionicotoxins, and batrachotoxins.
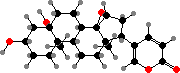
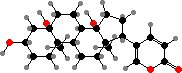
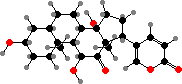
Most Bufonidae (toads) also produce bufadienolides. These are steroids with a strong effect on heart activity, like digitalis potentially fatal but in smaller amounts used in traditional medicine. Some more widespread examples are marinobufagin, telocinobufagin, and arenobufagin, found both in free form and as sulfates or organic esters.
Theropsida, the mammals and extinct relatives, are terrestrial or secondarily aquatic air-breathers. Living kinds typically have a covering of hair, differentiated teeth, and nurse their young with milk. Monotremes have amniotic eggs, which develop in air, but others all have live birth. Most of the larger animals belong here.
Mammals have good senses of smell and use diverse volatiles in communication. Some Carnivora, which often have sulfur compounds, also repel predators with sprays from anal glands. This is best developed in Mephitidae (skunks) with mainly 2-butene-1-thiol, 3-methylbutane-1-thiol, and often thioacetates that break down with water to release more.
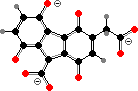
Skin and fur pigments here are mostly limited to melanins. Hippopotamus also produce alkaline secretions with a unique red pigment, hipposudoric acid, which exists in ionized form and otherwise degrades into brown polymers. This helps protect their skin from both ultraviolet and bacteria.
Sauropsida, the reptiles and birds, are largely terrestrial with amniotic eggs. They are distinguished by skeletal details but among living groups are more plainly set apart by body coverings of corneous scales or scutes. Birds also have scales modified into feathers, front limbs into wings, and many other adaptations allowing flight.
Birds have lost pterin pigments except in eyes, but carotenoids like astaxanthin, canthaxanthin, lutein, and zeaxanthin give skin or feather colours in various groups. Biliverdin and protoporphyrin are common in eggshells, and in Musophagidae (turacos) the feathers have red turacin or a green oxidized form, uroporphyrins with copper ions.
A few birds have also developed other unique feather pigments. Penguins have fluorescent yellow spheniscins of unknown structure. Parrots have simple polyenes called psittacofulvins, which take the form of aldehydes like octadecaoctaenal in red feathers and are converted to corresponding acids in yellow feathers.
Passerine birds often have carotenoids including some more unusual kinds. The most common are canary xanthophylls, keto-carotenoids where the chain of double bonds is cut like in tunaxanthins, shifting the colour to yellow. There are also some deeper reds like rhodoxanthin and even purples, particularly cotingin from Cotinga and relatives.
The annelids are a large group of mainly coelomate worms. Most have bodies divided into a short preoral region, sometimes part of a developed head, and a series of segments that usually have setae, though this plan has been lost in a few groups. Some also share ciliated larvae in the form of trochophores, also found in molluscs and other related groups.
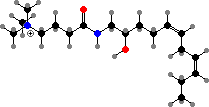
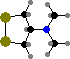

A few different annelids are toxic. Amphinomidae have protective bristles, found in Eurythoe fire worms to contain complanine, causing painful inflammation when they are handled. Some others have venoms or antimicrobial compounds. Lumbrineris are notable for producing mucus with nereistoxin, which is poisonous to animals like insects and fish.
Hemoglobins are common here, variously in cells or directly in blood, and give bright red colours to some worms. There are also some families with chlorocruorins, which have a variant of heme missing a carbon atom and are green when dilute, as well as a few with respiratory hemerythrins where iron ions are attached directly to the protein itself.
Related tetrapyrroles are also accumulated sometimes, for instance biliverdin conjugates in some green nereidids. In the echiurids Bonellia the larger females are coloured green by bonellin, a sex-determining pheromone that induces new larvae settling on them to instead become small dependent males.
Molluscs share a general body plan with a thickened dorsal mantle, which folds over a cavity around the anus and gills and often forms a calcareous shell, and muscular foot. The organs are concentrated as a visceral mass with an open circulatory system or hemocoel. Most also have a unique toothed radula adapted for various types of feeding.
Most shell pigments are melanins or uncertain polyenes. A few marine shells also show red fluorescence under ultraviolet from uroporphyrin, and though this often does not relate to visible colour its derivatives give pink or brown in at least some top snails. Bilins are known as shell pigments in the related turban snails and abalones.
Gastropoda, the snails and allies, are a large group found from ocean depths to deserts. Most are creeping types with a flat or coiled shell and develop by torsion, where one side is reduced and so the mantle cavity is turned to the front, but others are soft-bodied and secondarily streamlined. The gills too may be replaced, for instance by the whole cavity acting as a lung.
Many gastropods with reduced or no shells obtain protection from their food in the form of pigments or toxins. Nudibranchs most commonly store terpenes from dietary soft corals or sponges. Some also produce their own, like Dendrodoris with olepupuanes and derived polygodial, a hot-tasting dialdehyde known from several land plants and toxic to fish.

In addition various carotenoids are taken up for protective colouration, and aeolidids can obtain stinging cells from their prey. Several alkaloids from other animals are likewise echoed in nudibranchs and other carnivores: tambjamines in Tambja and Roboastra, trisoxazoles in Hexabranchus, bromotyrosines and quinolines in Tylodina, and kuanoniamines in Coriocella.
Sea hares like Aplysia store mainly terpenes and halogenated compounds from red algae and many sacoglossans from green algae, some also retaining chloroplasts. Others in these groups produce defensive polypropionates like elysione in Elysia, and so do a few other marine slugs and relatives, for instance predatory Clione with pteroenone.
Neogastropod snails sometimes use toxins to subdue prey, for instance peptide venoms in cone snails and relatives, and some whelks like Neptunea produce enough tetramethylammonium to also be poisonous if eaten. This acts on acetylcholine receptors, as do other choline esters like murexine from mucus glands of Muricidae (murex snails).
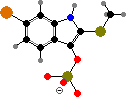
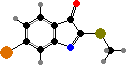
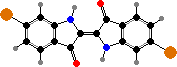
Muricids also produce indoles like tyrindoxyl sulfate, and in some the egg masses contain derived tyrindoleninone and its dimers as antimicrobials. Sunlight breaks these down into purple 6,6′-dibromoindigo and sulfides. Purple and related colours can also be made on fabric by fermenting dead or milking secretions from live snails and were important ancient dyes.
Arthropods are common in nearly every habitat. They make up one of several phyla that share chitinous cuticles, without cilia and periodically molted during growth, and in this case forming an exoskeleton with rigid sclerotized plates. These also all have variously reduced coeloms, here replaced as the main body cavity by a hemocoel.
The body is divided into segments with paired appendages, which are characteristically jointed. These are more or less specialized, with sensory antennae and mouthparts near the front, and various legs, gills, or so on further back. Matching this the segments are generally grouped into larger tagmata such as a head, thorax, and abdomen, or combinations of these.
Many visible colours here come from ommochromes, found as waste products but also typical light screens in compound eyes, and as the main pigments in some groups like spiders, isopods, libellulid dragonflies, and nymphalid butterflies. These are formed as dimers or oligomers of hydroxykynurenine, itself a light yellow pigment in for instance crab spiders.
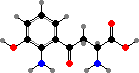
The main dimers are orange-yellow xanthommatin, red dihydroxanthommatin, and derivatives, which have been reported as the L- isomers in hornworms and decarboxy forms in libellulids. The oligomers are various sulfur-containing ommins, coloured black to brown, also typical of eyes and pigment cells in cephalopods and a few other animals.
Arachnida have bodies more or less divided into a cephalothorax and abdomen, the former with variously specialized chelicerae and pedipalps followed by four pairs of legs. There are no antennae or mandibles. Most are terrestrial, breathing with book lungs or tracheae and feeding on fluids or by liquefying prey.
Spiders, scorpions, and many pseudoscorpions ambush prey with venom. These are mostly peptides but there are also spiders with acylpolyamine neurotoxins, particularly orb weavers and some funnel spiders, such as argiotoxins in Argiope. Among non-venomous relatives the whip-scorpions are notable for spraying acetic acid when threatened.
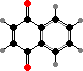

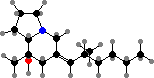
Harvestmen have scent glands with repellent compounds, variously carried by regurgitated fluid in channels along the body, sprayed, or dabbed on legs. These include 1,4-naphthoquinone, as in Phalangium, and acyclic ketones, as in sclerosomatids. Laniatores have types like 2,3-dimethylbenzoquinone in most gonyleptoids or 5-ethyl-2-methylphenol in other relatives.
There are also oil glands in many oribatid and astigmatid mites used for defense or for pheromones. The most common components are hydrocarbons, geranial and neral, γ-acaridial, and their derivatives. Among the others are some alkaloids, for instance pumiliotoxins from Scheloribates, also known in ants and taken up as a convulsive toxin by some frogs.
Diplopoda, the millipedes, have a distinct head and trunk, which is mostly undifferentiated except the first four segments, including a legless collar. The remaining body segments are all characteristically united in pairs, with a calcified dorsal plate over every four legs. The head has one pair of antennae, mandibles, and maxillae. All are terrestrial and most feed on decaying plant material.

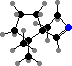
Some orders of millipedes defend themselves primarily by rolling up into a ball, among which the glomerids also have dorsal glands that release sticky fluid. This is partly mechanical protection but also has bitter glomerins, which paralyze some predators like spiders. Most other orders are elongate and have lateral defensive glands with a variety of different substances.
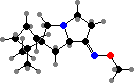
Alkaloids are typical of Polyzoniida, for instance polyzonimine in Petaserpes and spiropyrrolizidine 236 in Rhinotus and some frogs. Spirostreptida, Spirobolida, and Julida form a group that release or spray methyl-p-benzoquinone and related quinones, also found among beetles and other insects, noted for staining skin and noxious to many different animals.
The large order Polydesmida are more variable but most can release hydrogen cyanide. This is formed in bicompartmental glands by breaking down mandelonitrile, reported as the D- isomer in Harpaphe, and in some also benzoyl cyanide. Both these precursors also occur among geophilomorph centipedes, and mandelonitrile again in a few different insects.
Ostracoda are one of several classes of crustaceans, which generally have two pairs of antennae, branched appendages, and are largely aquatic. Many have a heavy carapace; ostracods have one divided into two valves that enclose the body and limbs, of which there are six or fewer behind the head. They vary from minute benthic filter-feeders to planktonic predators.
The scavengers Conchoecia are among various crustaceans with bioluminescence from coelenterazine in body glands. This has an imidazolinone ring derived from amino acids, which is broken to give a peroxide that emits blue light. So far its synthesis is confirmed in copepods and shrimp, though it is also taken up for light production in many different marine animals.
Many Cypridinidae instead make luminous sprays, used to escape predators and in some like Photeros for male courtship displays. The cypridinid luciferin, or vargulin, also has an imidazolinone ring but from different amino acids. Here it is expelled from labral glands along with an oxidizing enzyme. It is also employed by a few predatory fish like pempherids and midshipman fish.
Malacostraca are diverse crustaceans with both thoracic and abdominal appendages. The thorax has 8 segments, often one or more with maxillipeds for feeding and the others with legs, and the abdomen has 6 or 7 segments with paddle-like limbs. Many have a carapace and the eyes are often on stalks. Most of the larger crustaceans belong here.
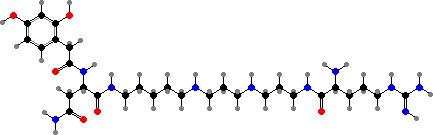
A few kinds are terrestrial, notably some isopods or woodlice. Like glomerids many can defend themselves by curling up or releasing sticky secretions, which in Porcellio were found to deter spiders with quinolines like the one shown. They mainly have ommins as colouring, but astaxanthin-protein complexes are typical of other orders like decapods.
Insecta are derived from crustaceans but largely terrestrial. All share a basic body plan divided into a head with one pair of antennae, thorax with six legs, and abdomen, which has gills in some aquatic larvae but otherwise has only limited sensory or reproductive appendages near the end. Breathing takes place through spiracles and so is not dependent on moisture.
The appendages are not branched, but in most adults the second and third thoracic segments have pairs of both legs and wings, which are either membranous or modified but in general allow flight. The winged insects have adapted to nearly every terrestrial habitat, and are also common in freshwater, with more described species than all other living things.
Insect pigments often include melanins and carotenoids, with yellows like lutein commonly mixed with protein-bound biliverdin to give grass-green colours. Pterins are also widespread, found alongside ommochromes in compound eyes and like them sometimes as body or wing pigments in different groups.
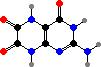
Typical pterins here include red erythropterin, yellow xanthopterin, and leucopterin, which is colourless to us but absorbs ultraviolet so is visible to most insects. These for instance occur in many heteropteran bugs, vespid wasps, pierid butterflies, and also as light-scattering grains giving complementary green to blue colours in pond damselflies.
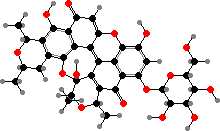
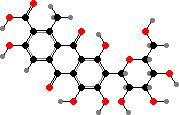
Sternorrhynch bugs often have quinone pigments. In aphids these are largely dimers, for instance green viridaphin A1 glucoside in Megoura crassicauda. Scale insects have red anthraquinones that deter some predators like ants, notably carminic acid from Dactylopius, which is used as the dye and food colouring cochineal.
Biliverdin in most insects is the same type as other groups, called IXα, but many caterpillars an alternate form called IXγ or pterobilin where the heme ring is broken in a different place. This and its derivatives sometimes also give colour to adult wing membranes exposed as bluish patches, notably among swallowtail butterflies like Graphium.
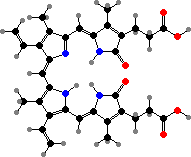
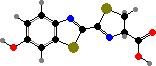
A few different groups include some bioluminescent types, but most notably elateroid beetles: fireflies, glowworms, and some click beetles. Light is produced by both adults and larvae, serving to attract either prey or mates. The colour varies from red to green, the result of different oxidizing enzymes, but sharing lampyrine as the luciferin.