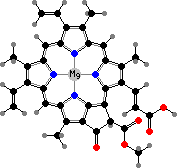
Chlorophyll c2
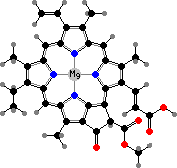
Chlorophyll c1
Introduction -
Common -
Bacteria -
Plantae -
Chromista -
Protozoa -
Fungi -
Animalia -
References
Cryptista -
Ciliophora -
Miozoa -
Ochrophyta
Chromists vary from single cells to multi-cellular seaweeds. Several major lineages have chloroplasts, originally taken from red algae but with different pigments and characteristically surrounded by one or two additional membranes. Others feed by absorption or ingestion, often with thread- or needle-like pseudopodia, including many traditionally considered protozoans.
Photosynthetic chromists usually have chlorophyll c2 in addition to a, and some also have chlorophyll c1, which has an ethyl in place of one vinyl group. These are unique in occurring as free acids, rather than esters with a non-polar side chain. Phycobiliproteins are found in cryptomonads but have been lost in all others.
Instead these groups have allenic or acetylenic carotenoids, often in large amounts that give them a golden or brownish colour. The most important of these types is fucoxanthin, which is the main pigment in haptophytes, golden algae, brown algae, and diatoms. Most also have smaller amounts of diadinoxanthin, as do the unrelated euglenid flagellates.
Cryptista are mostly single cells with two flagella and characteristic ejectisomes. The principal group are the cryptomonads, where both occur around a groove or pocket set behind the front. Most kinds are photosynthetic with one or two plastids, which unlike in other chromists contain starch grains and a degenerate nucleus retained from the ancestral red alga.

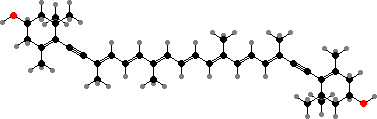
As noted, these also retain phycobiliproteins in addition to chlorophyll a and c2, which variously colour them red, green, or bluish. They are also unusual in lacking epoxy carotenoids like diadinoxanthin; instead the main type is alloxanthin, together with smaller amounts of other diacetylenic carotenoids.
Ciliates take the form of single cells or simple colonies. They are unique in having smaller and larger nuclei, the former diploid with normal mitotic division, the latter polyploid and simply pinching in two. They lack plastids and generally ingest food through a distinct mouth. Most also have many short flagella, called cilia, arranged in rows or fused into compound structures.
The membrane is also associated with a layer of packed vesicles or alveoli, and often various types of extrusomes that expel material, for instance toxicysts to paralyze prey or trichocysts to block predators. Cortical granules found in karyorelictids, heterotrichs, and spirotrichs may have pigments or other chemical defenses; most other ciliates are only coloured by food or algae.
Heterotrichea typically have body cilia in rows, arising from paired bases with a characteristic arrangement of microtubular ribbons. The oral cilia on the other hand form a series of compound membranelles, which run from the front of the cell and clockwise into its mouth, used for both feeding and movement.
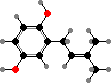
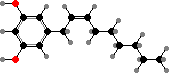
Extrusive granules are found throughout this group and release various compounds that deter predators. For instance Spirostomum ambiguum, among the largest ciliates, defend themselves with mono-prenyl hydroquinone. The omnivores Climacostomum virens have climacostol, which might also be used in capturing prey ciliates.
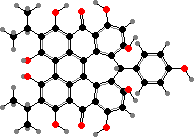

In still others the granules have quinone dimers with various colours, such as blepharismins from red Blepharisma and stentorin from the blue trumpets Stentor coeruleus. These act both as photosensitizers to repel predators and receptors for light avoidance, though in too bright light can become toxic to the heterotrichs themselves.
Spirotrichea have a series of oral membranelles like in heterotrichs, but the other cilia do not have the same arrangement. In most the latter are reduced and sometimes absent, particularly in planktonic types, or alternatively fused into prominent compound tufts called cirri, which are mainly used for crawling on surfaces.
Various terpenoids have been found in marine Euplotes. These are mainly defenses specific to regional varieties, such as rarisetenolide epimers in different E. rariseta, but euplotins common to E. crassus instead act as contact toxins against competing species. Pigment granules occur in a few other spirotrichs, for instance Pseudokeronopsis rubra with red keronopsins.
Miozoa are a diverse group of flagellate or parasitic cells, related to ciliates and with similar alveoli though generally haploid. Many have an apical structure adapted for sucking fluids from or entering other cells, but this has been lost in many free-living types. Plastids are found in most lineages although photosynthesis is almost limited to dinoflagellates.
The dinoflagellates have peculiar nucleoproteins and outside some small colourless groups, these go with a distinctive sort of nucleus where the chromosomes are attached to the membrane. Most types have two flagella set in grooves, one circling the cell and one directed to its posterior, though this varies and a few have non-motile or colonial forms.
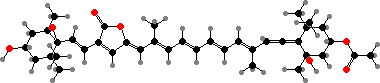
Photosynthetic dinoflagellates are among the most common planktonic algae. The main carotenoid is usually peridinin, a unique kind formed by the loss of a C3 unit. In a few genera, though, the chloroplasts have been secondarily replaced by ones from haptophytes or green algae, which then contain fucoxanthin or other pigments.
Most ochrophytes have chloroplasts, though some also capture other organisms, and generally depend on plastids for lipid synthesis. They make up part of the diverse heterokont lineage, which vary greatly in form and habitat but typically share spores, gametes, or other motile cells with one or two unequal flagella, of which the anterior one has distinctive tripartite bristles.
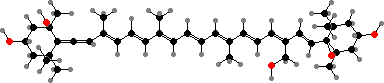
Fucoxanthin is the typical pigment but has been lost in a few largely freshwater lines with green chloroplasts. For xanthophytes (yellow-green algae) the main carotenoids are usually diadinoxanthin and esters of vaucheriaxanthin. The latter also occur in others like eustigmatophytes, with mainly epoxides of zeaxanthin and only chlorophyll a.
Phaeophyceae, the brown algae, have cell walls with fine channels similar to many plants. They are primarily marine and vary from simple filaments to giant seaweeds, often differentiated into leaf-like blades, stems, and holdfasts. In most there are separate diploid and haploid individuals, though in more complex forms the latter are minute or reduced.

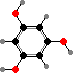
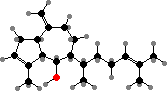
Brown algae have vesicles called physodes with phlorotannins, unique protein-binding polymers made from phloroglucinol units. These deter herbivores and colonizing organisms, along with other functions like absorbing ultraviolet. Some also have C20 terpenoids, for instance Dictyota with pachydictyols among others, which likewise protect them from feeding and settling.
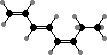
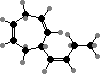
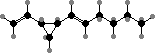
A series of C8 and C11 hydrocarbons are found throughout this group and contribute part of the ocean’s characteristic scent. These mainly act as pheromones that one gamete, usually larger, releases to attract the other; fucoserratene in Fucus and ectocarpene in Ectocarpus are representative. Dictyopterenes from Dictyopteris also serve to inhibit some herbivorous fish.