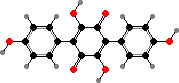
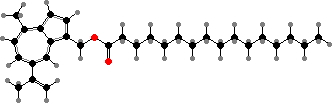
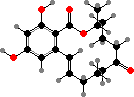
Atromentin
Introduction -
Common -
Bacteria -
Plantae -
Chromista -
Protozoa -
Fungi -
Animalia -
References
Agaricomycetes -
Pezizomycetes -
Leotiomycetes -
Sordariomycetes -
Eurotiomycetes -
Lecanoromycetes -
Arthoniomycetes
Fungi have cell walls and mainly live as saprobes, parasites, or in other symbiotic relationships. Flagella are usually absent except in spores of some simple types, which grow into enlarged cells with rhizoids. Most others form masses of filaments called hyphae that penetrate and absorb nutrients from the medium, though there are also some like yeasts that have reverted back to single cells.
These are part of the Basidiomycota, which during sexual reproduction form spores on the outside of cells called basidia. These typically develop into septate hyphae, which fuse to form a dikaryon containing a mix of nuclei from two or more parents. This is usually the main stage; for the most part diploid nuclei only occur during spore formation.
Agaricomycetes usually form large fruiting bodies like mushrooms. They mainly include types with club-shaped and non-septate basidia, though some are exceptions, often on external structures like spines, tubes, or lamellae. They are most notable as saprobes and for mycorrhizal associations with plants, where they provide water and minerals to roots in exchange for sugars.
A variety of pigments are found here, notably carotenoids and terphenyls. The most common of the former are β- and γ-carotene, also typical of related classes like jelly fungi. Canthaxanthin is typical of some bright orange Cantharellus (chanterelles), while Clathrus and several other stinkhorns have red colours from lycopene.
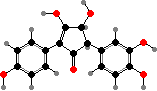
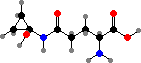
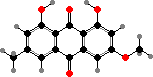
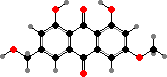
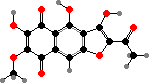
Terphenyls and their derivatives are a broad group of compounds derived as dimers of amino acids, usually tyrosine, which gives rise to atromentin and through it most others. Atromentin is itself accumulated as a brown pigment in some types and causes a violet colour change when they are treated with alkali.
Thelephorales and Polyporales are united mainly by genetic trees. These include many of the mushrooms with basidia on spine-like projections, and polypores, where they are in tubes on the underside of a tough fruiting body with an eccentric or no stalk. Some also form smooth crusts on wood, and a few are agarics, with a central stalk and basidia on distinct lamellae.

Terphenyls are common pigments in both these groups. Atromentin is found in Thelephorales such as Hydnellum aurantiacum, which get their colour from it and its red dibenzoate aurantiacin. Thelephoric acid, a widespread violet derivative, is also a typical component in this order and causes the flesh to turn blue or green in alkali.
Polyporales also have thelephoric acid or related compounds on occasion, for instance bright blue Terana caerulea with pigments derived from similar hydroquinones. Several others have phenylalanine-derived terphenyls, notably brown polyporic acid in genera like Hapalopilus, where it can make up as much as 40% of their dry weight.
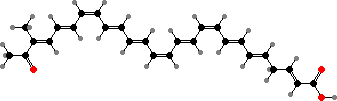
Still other genera have entirely different pigments. The edible Laetiporus sulphureus (sulfur polypores) get their orange-yellow colour from laetiporic acid and related polyenes like the one shown. Another type occurs in Pycnoporus, which have red-orange cinnabarin and other phenoxazines.
Russulales are another genetic group with some polypores, spined fungi, and agarics, though they usually share some features like spore ornamentation. Special hyphae and cells with resinous or granular contents are very common here; they are best known in Lactarius (milk caps), which exude milky latex when injured.
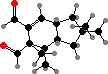
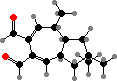
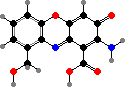
These produce a variety of C15 terpenoids, often in the form of esters that are broken down as an activated defense in damaged tissue. The most common is stearoylvelutinal, which gives rise to different pungent dialdehydes like isovelleral and velleral from L. vellereus, L. rufus, and various others. This serves to deter both herbivores and microbes.

Milk caps with coloured juice are generally edible when cooked. L. indigo gets its deep blue colour from stearoyldeterrol, and a reduced form is the orange in relatives like L. deliciosus. These break down to give purple lactaroviolin, and in the latter also blues like lactarazulene and yellows, so that they develop green bruises.
Boletales make up the boletes and their allies, most of which live in mycorrhizal associations with trees. The fruiting bodies in boletes are stalked, fleshy mushrooms where spores are discharged from tubes rather than ridges or lamellae. Some agarics and false truffles also belong here, as well as the dark sack-like earth-balls.

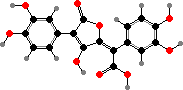

The main pigments here are terphenyl derivatives, especially pulvinic acids. Variegatic acid and its air-oxidized form variegatorubin are common types responsible for the yellowish and red colours in many genera. Some Boletus and Suillus also oxidize these acids to quinone methides when cut or bruised, turning the flesh bluish.
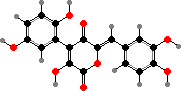
Several other genera turn bluish or brownish on injury from oxidation of cyclopentanoid derivatives, for instance colourless involutin in Paxillus involutus (brown roll-rims). Besides these types many Suillus have orange-red grevillins, formed by a related but different pathway from terphenyls, and some others have pigments derived from quinones or phenols.
Agaricales make up the majority of agarics, including for instance those where the lamellae are separated from the stalk. Among the Agaricineae there are also some more unusual families like puffballs, with internal spores released by rupturing, and bird’s nest fungi, which form small cups with spore chambers scattered by rain.
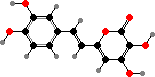
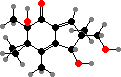
A number of Mycena and other Marasmiineae are bioluminescent, giving off green light by oxidation of 3-hydroxyhispidin. In some this attracts insects to carry spores but it can also occur on substrate hyphae. Omphalotus in this group are also notable for terpenoids like illudin S, which causes gastrointestinal poisoning, as well as terphenyls and pulvinic acids.
Pigments related to the betalains of caryophyllid plants are found in red to yellow species of Hygrocybe and Amanita. In the former these are hygroaurins, which have yellow muscaflavin as a precursor in place of betalamic acid. Muscaflavin also occurs in A. phalloides (death caps), one of several species with extremely deadly peptide toxins.

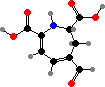
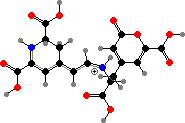
Other Amanita like A. muscaria (fly agarics) and edible A. caesarea have red to orange caps that also have true betalains. These include some of the same yellow betaxanthins found in plants along with orange muscaurin II, derived from the unusual amino acid stizolobic acid, and purple muscapurpurin of incompletely known structure.
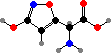
A. muscaria were some of the first known hallucinogenic mushrooms, with ceremonial use by Siberian tribes. The main compound involved is ibotenic acid, which occurs with its derivatives muscaaurin I and muscimol; the last is the active form triggering GABA receptors, resulting in symptoms from nausea to hallucinations and convulsions.
Anthraquinones and dihydroanthrones, including a number of types found in other fungi and plants, are common pigments in Tricholoma and Cortinarius. These cause red or violet colour changes with alkali. The quinones are largely acids like orange endocrocin, or red dermorubin in the latter genus, but others like physcion also occur.
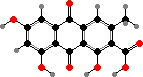
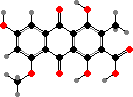
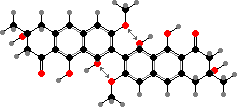
The dihydroanthrones mostly occur in a variety of dimers, of which flavomannin dimethyl ether and derivatives are the most common. In these rotation of the central bond is blocked and so they occur as separate atropisomers. B isomers are the main pigments of yellow T. sulphureum, while some Cortinarius have only A isomers; still other species have a mix of both.
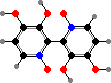
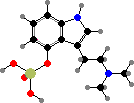
There are also a few Cortinarius with orellanine, a strong toxin causing kidney failure. Other Agaricineae with notable compounds include Psilocybe and other genera with psilocybin, a hallucinogen with traditional use in North and South America, and Coprinopsis with coprine, which is non-toxic on its own but blocks detoxification of alcohol.
The remaining classes of fungi here are all Ascomycota. These variously take the form of single cells or septate hyphae and have a dikaryotic stage similar to Basidiomycota. This though is typically limited to fruiting bodies and instead of basidia, sexual reproduction takes place by the production of spores on the inside of cells called asci.
Pezizomycetes usually have fruiting bodies in the form of apothecia, which develop an upper layer of exposed asci, and here these in turn expel spores through an operculum or lid. They vary from cup-like to larger stalked forms with wrinkles or pits. Some types like truffles are exceptions, remaining closed underground and depending on animals to distribute spores.
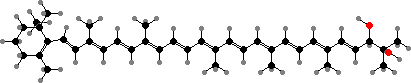
Many cup fungi are brightly coloured with carotenoid pigments. Besides common types like γ- and β-carotene these include some more unusual sorts, particularly ones with hydroxylated end groups like plectaniaxanthin. This is for instance the main colour in Sarcoscypha coccinea (scarlet cup fungi), mostly as fatty acid esters.
Leotiomycetes usually have smaller apothecia where the asci open through pores, found as cup- to club-shaped bodies on decomposing material, soil, or plants. The parasitic powdery mildews however instead have cleistothecia, some with radiating appendages, which remain closed and typically release spores when they are burst by moisture.

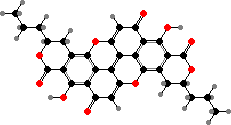
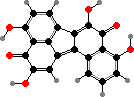
This is another group where β-carotene is a common pigment. There are also a few with quinone dimers, notably xylindein in Chlorociboria, which gives the cups a distinctive bluish-green colour as well as staining the wood where they grow. Some others are related to melanin precursors, for instance bulgarhodin in dark Bulgaria.
Sordariomycetes also have asci that open through pores but typically produce them in minute perithecia, which are enclosed except for a small opening or ostiole. Here these develop through the formation of distinct walls around the dikaryon. A few have lost the ostioles, though, and in others reproduction is mainly or entirely through asexual spores called conidia.
Hypocreales have colourless ascospores and usually light or coloured perithecia. In asexual forms conidia are typically produced on the ends of flask-shaped cells called phialides. This group includes a number of common molds and endophytes or parasites of plants, other fungi, and insects.
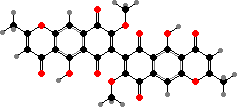
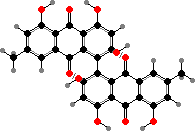
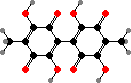
Quinones and dimers are common here. Fusarium produce red naphthoquinones like aurofusarin, also in some Hypomyces. Others have anthraquinones like orange skyrin in edible H. lactifluorum, an emodin dimer found in several groups of fungi. Parasitic Hyperdermium also have skyrin and Beauveria have oosporein, both toxic to their insect hosts.
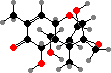
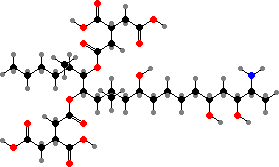
Several genera have trichothecene toxins that interfere with protein synthesis in plants, animals, and other fungi, for instance deoxynivalenol or vomitoxin from Fusarium like F. graminearum on grain. Some of the same species also have zearalenone, which has estrogenic effects, while others like F. fujikuroi have fumonisins responsible for various animal diseases.
Xylariales commonly have walled perithecia embedded in masses of fungal tissue called stromata, which vary from simple growths to branched stalks. The asci are persistent and occur among sterile filaments called paraphyses. Most kinds are saprobes on dead plant materials like wood or sometimes parasites of living plants.
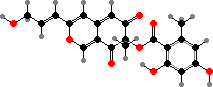
Stromata in Hypoxylon and relatives are often coloured and are notable for azaphilone pigments, named for their tendency to react with amines. The most common are orange-red mitorubrinol and variations, for instance in H. fragiforme on beech. There are also various binaphthyl derivatives related to melanins.
Eurotiomycetes have asci that are either fissitunicate, with a functionally separate inner wall that expands and bursts through the outer one to expel the spores, or have lost the outer portion and simply disintegrate in place. The first occur in perithecia while the second are often found in cleistothecia or powdery mazaedia. Asexual forms are also common.
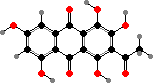
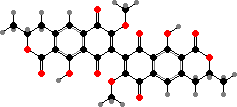
Many of the perithecial types form lichens like in the following classes, and a few have cortical pigments like red draculone in Pyrenula cruenta. Then some other groups here include animal pathogens, among which the quinone dimer xanthomegnin is typical of the skin fungi Trichophyton as well as one of many toxins found in Eurotiales.
Eurotiales produce round asci inside cleistothecia, both breaking down to release the spores, or sometimes on unenclosed hyphae. Distribution is typically by conidia formed in chains on phialides like in Hypocreales. Most are saprobes, well known as molds on spoiled foods and some products like blue cheeses.
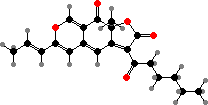
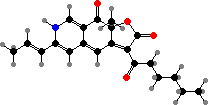
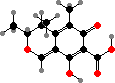
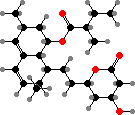
Azaphilone pigments are common in Talaromyces and Monascus, for example orange rubropunctatin and its purple-red derivative rubropunctamine. Species like M. ruber are used in making red yeast rice but can also produce citrinin, a kidney toxin, as well as lovastatin, which is used as a drug to inhibit cholesterol production.
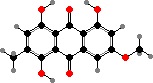
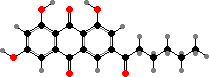
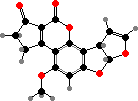
Pigments here also include anthraquinones like skyrin in Talaromyces and red erythroglaucin in a few Aspergillus. Some others like A. flavus have norsolorinic acid and derivatives, which ultimately serve as precursors for colourless aflatoxins. These are notoriously strong carcinogens causing liver damage and so can be dangerous contaminants in food.
This is the largest group of fungi that form lichens, capturing and enclosing particular kinds of green algae or sometimes cyanobacteria that then provide them food through photosynthesis. Most are able to live with little other nourishment or moisture and take the form of thalluses with small but persistent fruiting bodies.
Lecanoromycetes usually have apothecia and produce asci with a thick apex, which opens through a combination of its inner wall elongating and outer wall breaking. A handful have perithecia or else mazaedia where spores are released as a powdery mass. Nearly all grow exclusively as lichens, found in a wide range of terrestrial habitats.
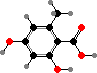
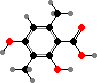
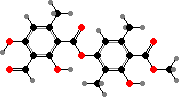
Many lichens produce different esters from orsellinic or β-orsellinic acids, called depsides and depsidones, as crystals on the hyphae. The most common here is atranorin, which is found in the upper cortex and absorbs ultraviolet; in some people it is a contact allergen or photosensitizer. This and various other cortical compounds vary with light and so screen the algae.
Others are found on interior hyphae. Some more common examples are gyrophoric acid, a plant growth inhibitor; norstictic acid, which binds to metal ions; and stictic acid, another allergen. They also inhibit some bacteria. These are colourless, but many of the aldehydes show as yellow or red in alkali, and orsellinic acid derivatives as red in hypochlorite.
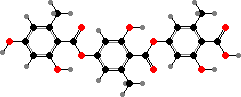
Various lichen dyes are based on similar changes. Lichens like Umbilicaria and Ochrolechia with gyrophoric or sometimes lecanoric acid can be used to make dyes similar to orchil. Other types with large amounts of aldehydes, like Lobaria with norstictic and stictic acid, give fast yellow or brown colours when boiled with wool and other fibres.
Brighter pigments that occur here are mostly pulvinic acids or quinones. The former are typically yellows like rhizocarpic acid, found in various types like Acarospora and Rhizocarpon, and calycin, found in Pseudocyphellaria as well as other lichens like Candelariales. The latter are more variable and give more orangish yellow to red colours.
Teloschistales are orange to yellow lichens with crustose, foliose, or occasionally stalked thalluses. The spores are typically colourless and polarilocular, with two cells divided by a thick median wall that confines them to the ends though often has a canal between them.

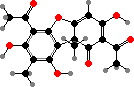
The characteristic colour comes from anthraquinones, especially physcion, also found in various other fungi and plants. Derivatives like fallacinol are also common. Here these act as light screens, with larger amounts made in response to exposure, along with other protective effects like deterring feeding by snails.
Lecanorales are a diverse group. The apothecia are usually round with colourless spores, though some have mazaedia with a mass of dark spores instead. In those categories most of the larger and non-cyanobacterial fruticose lichens, which is to say ones with erect or pendulous stalks, belong here along with a mix of crustose, scale-like, and foliose relatives.
The main light screens here atranorin and usnic acid, which occurs as both the (+) and (–) enantiomers in different species. These give light yellow colours to many more sun-exposed lichens. They also show activity against a variety of bacteria, plants, herbivores, and so on, and have been used as antiseptics but again can be strong contact allergens.
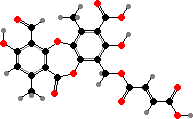
Fumarprotocetraric acid is also relatively common here, for example in many large boreal Cladonia and Cetraria; this bitter depsidone makes them less palatable to grazers like reindeer. Some Cladonia also form rhodocladonic acid, for instance as a response to mites, a quinone which colours the apothecia pink to bright red.
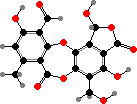
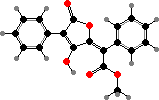
Many species in the family Parmeliaceae have alternative depsidones like salazinic acid, including some used for dyes. This group also includes a few toxic lichens with vulpinic acid, which has been used for both dyes and animal poisons; this is the bright yellow pigment in Letharia and Vulpicida, poisonous to insects and snails as well as carnivores.
Arthoniomycetes mostly form lichens with green algae. The asci tend to be somewhat fissitunicate, though the walls may not entirely separate, and surrounded by extensions of generative tissue. They usually occur in round to elongate apothecia, often in the form of more peculiar slit-like or branched hysterothecia, though this varies.
Many kinds have compounds of the same types as Lecanoromycetes. These include pulvinic acids like calycin and vulpinic acid in Chrysothrix, quinones like red chiodectonic acid in Herpothallon, and various depsides and depsidones. The most common here is lecanoric acid, a plant growth inhibitor similar to gyrophoric acid.
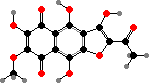
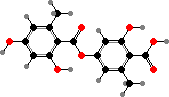
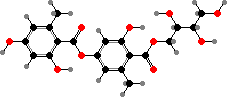

A number of types also have erythrin, a lecanoric acid ester, identified as the (+) enantiomer in at least Roccella montagnei. This is one of several species used for orchil and litmus, phenoxazine dyes made by fermenting them with ammonia. The resulting colour varies from red in acid to blue in alkali and has long been used as an indicator for them.