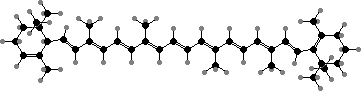
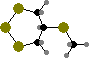
α-Carotene
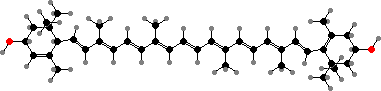
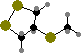
Lutein
Introduction -
Common -
Bacteria -
Plantae -
Chromista -
Protozoa -
Fungi -
Animalia -
References
Rhodophyta -
Chlorophyta -
Marchantiophyta -
Bryophyta -
Tracheophyta:
Angiospermae
The remaining kingdoms are all eukaryotes, with typically larger cells including a complex system of protein filaments and internal membranes. These include a special compartment for the DNA, the cell nucleus, which is divided into several chromosomes. Division generally takes place by mitosis, where two spindles of microtubules allocate the copies to each new nucleus.
Many eukaryotes also have some form of sexual reproduction involving combination of genes from two individuals. This usually involves an alternation between haploid cells with one copy of each chromosome and diploid cells with two. New diploids are formed by fusion of gametes, and new haploids by meiosis, a sort of abridged mitosis. Either stage may dominate the life-cycle.
Eukaryotes also have organelles originally derived from an enclosed proteobacteria, usually in the form of mitochondria, which have two membranes and are involved in aerobic respiration. Plants and some other algae also have plastids, derived from cyanobacteria, most often in the form of chloroplasts which conduct oxygen-producing photosynthesis.
True plants range from minute cells to giant trees. They are set apart by having plastids with only two membranes, even in the few non-photosynthetic kinds, since they rely on the starch plastids produce as a food store. Most have cell walls and tend to be non-motile in the main vegetative stage, although many have at least some flagellate cells.
The photosynthetic pigments are still chlorophyll a and phycobiliproteins in glaucophytes and red algae, but the latter have been replaced in green plants. The carotenoids also vary but usually include β-carotene, zeaxanthin, and two more where one double bond is shifted, α-carotene and lutein. The last of these appears not to occur in bacteria or other algae.
Red algae are mainly marine and vary from single cells to complex seaweeds. The chloroplasts are usually red in kinds from deep water, but in those from coasts or freshwater they are often greenish, bluish, or dark brown. No flagella occur in the group, and they generally live on rocks or other surfaces, relying on currents to distribute their gametes and spores.
Florideophyceae include some branched filaments and algae with multiple layers of cells, which have multiple nuclei and are linked by pit connections. Most have similar haploid and diploid stages that produce gametes and tetraspores, respectively, but there is also a characteristic carposporophyte stage after the gametes fuse, which is typically filamentous and remains attached to its parent.

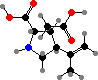
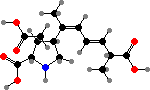
The neurotoxin α-kainic acid, which excites glutamate receptors, is found in Digenea simplex and a few other species, and its analogue domoic acid is found in Chondria armata. The latter is also known from some diatoms as a shellfish poison, but the concentrations here are lower and some of these seaweeds have been used as medicine to kill intestinal worms.
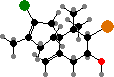
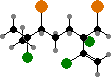
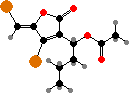
In many orders halogen compounds are common as defenses from both herbivores and colonizing organisms. These are usually terpenoids like elatol and others from Laurencia, and halomon, a potential antitumor agent from Portieria. Another notable type are fimbrolides from Delisea pulchra, which protect them from biofilms by interfering with bacterial signaling.
Green algae vary from single cells and simple filaments to differentiated types like stoneworts, and are in turn ancestral to other green plants. They usually have bright green chloroplasts with one or more pyrenoids and occur in many aquatic habitats, especially freshwater. Gametes and spores usually have two or more anterior flagella, which move in coordinated strokes.
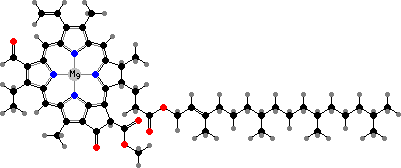
In place of phycobiliproteins these have a mix of chlorophyll a and b, which has one oxygen in place of two hydrogen atoms. These are bluish- and yellowish-green, combining to give the usual bright colour. The main carotenoid is most often lutein, along with β-carotene and epoxy derivatives of zeaxanthin. These are also the typical pigments of leaves in other green plants.
Ulvophyceae are set apart by motile cells with flagella inserted counter-clockwise and mitosis with persistent spindles. Most marine green algae belong here. They include various filaments and sheets of cells as well as many multi-nucleate kinds, which culminate in large siphonous forms similar to other seaweeds but not divided into separate cells.

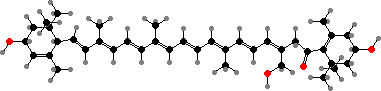
In many of these larger forms lutein is replaced as the main carotenoid by red-orange siphonaxanthin and its fatty acid esters. These help absorb higher frequency light, which is better able to penetrate into deep seas, and so play a role analogous to phycoerythrobilin in red algae from these habitats.
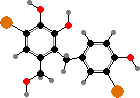
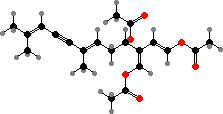
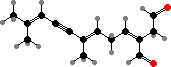
A few types have halogenated defense compounds like avrainvilleol in Avrainvillea. However, most instead have terpenoid acetates, which are broken down into toxic dialdehydes in damaged cells. For instance caulerpenyne from some Caulpera like C. taxifolia, an invasive species in the Mediterranean, forms oxytoxins that help render them inedible to many herbivores.
Charophyceae, the stoneworts, are multicellular weeds found in freshwater. The main stalks show apical growth, with whorls of short branches at intervals, and are anchored by a system of rhizoids. The cell walls are often calcified. This main stage is haploid, called a gametophyte, producing coiled sperm and eggs in special enclosed structures at the nodes.
These are closely related to terrestrial plants. They do not have a diploid sporophyte stage, as fertilized zygotes divide directly into new haploid filaments, but are similar to liverworts and mosses in many other respects. This includes the presence of flavones, found here as C-glycosyl derivatives in Nitella.

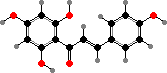
The other common genus is Chara from mainly calcium-rich water. These have a musky or garlic odour from sulfur compounds, which are thought to help keep them relatively free from epiphytes. For instance C. globulus produce charatoxin II, shown to inhibit photosynthesis in diatoms, as well as I which is more toxic to insects.
The remaining groups of plants are all multicellular and usually terrestrial. Hornworts, liverworts, and mosses are relatively small, without specialized vascular tissue and anchored by simple rhizoids. The main stages are gametophytes, which produce gametes in enclosed structures. A fertilized zygote develops into a diploid sporophyte, which remains small and dependent on its parent.
Liverworts themselves are more or less flattened with unicellular rhizoids. Some take the form of branching thalli, while others have simple leaves arranged in series on either side of the stem and sometimes below it. These are generally a single cell thick with no midrib. The sporophytes are short-lived, and there is normally no extensive filamentous stage as in mosses.
Many land plants have flavonoids, a diverse group of compounds with roles as sunscreens, defenses, pigments, signals, or so on, which share tetrahydroxychalcone as a common though unstable precursor. In liverworts and mosses they are mostly represented by flavones like apigenin and luteolin, found as C-glycosyl derivatives or O-glycosides.
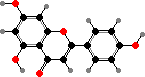
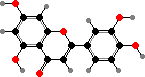
Such flavones have strong ultraviolet absorption though little visible colour. Some of these plants also have more apparent red flavonoids, especially in more sun exposed habitats. Riccionidin A, a flavylium ion found in cell walls, is the main type from several genera like Ricciocarpos; in still others the wall pigments are apparently polymers.
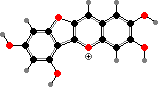
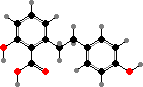

Most liverworts have cells with unique oil bodies, bound by a single membrane, that contain various protective compounds. A major group are bis-bibenzyls, unusual dimers related to lunularic acid, which is itself widespread and presumably acts as a hormone limiting growth. Marchantin A from Marchantia is an example, active against many bacteria and fungi.
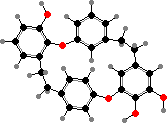
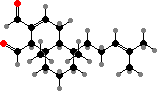
Many different terpenoids are also found in oil bodies as well as species without them. An example from a few different families is sacculatal in Pellia, Riccia, and others, one of several hot-tasting dialdehydes that deter herbivores. Various others are bitter, toxic, antimicrobial, or so on, while volatiles like pinenes are also typical in many liverwort scents.
Mosses usually have leaves in several ranks around the stem, often with midribs that are more than one cell thick. They develop from a branched filamentous stage, and the rhizoids likewise take the form of multicellular filaments. The sporophytes are photosynthetic until they mature, at which point the surrounding capsules generally open by means of a distinct lid.
The reddish pigments typical of some mosses are deoxyanthocyanidins, related to flavones but with a flavylium ion structure giving them visible colour. These often occur in cell walls, notably the red-violet sphagnorubins in Sphagnum (peat mosses). The simpler orange type luteolinidin is found as glycosides in some red Bryum species as well as a few ferns.
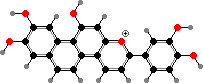
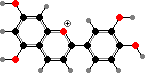
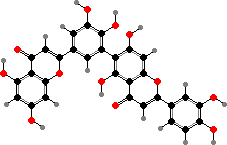
Many mosses also have lighter flavonoids in cell walls, where they mostly take the form of dimers like biluteolins instead of glycosides. The larger number of phenol groups helps these bind to proteins and high concentrations here are thought to help defend against fungi and insects as well as ultraviolet.
Vascular plants have reduced gametophytes and the main stages are instead larger sporophytes. These characteristically have a protective waxy cuticle and specialized transport tissues, xylem for distributing water and phloem for distributing sugars. Most are well-developed with distinct stems, roots, and veined leaves, though there are exceptions for each.
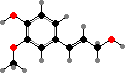
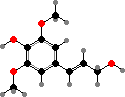
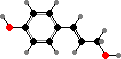
Cell walls in xylem are strengthened with lignin, which allows plants to stand upright, and it is also the main component forming wood in the cortex and bark of shrubs and trees. This is a complex irregular polymer formed mainly from coniferyl alcohol and sometimes sinapyl alcohol, particularly in flowering plants, as well as some p-coumaryl alcohol.
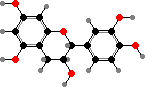
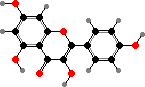
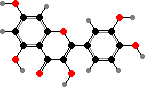
Many ferns and seed plants also have polymeric flavonoids, called proanthocyanidins or condensed tannins, as astringent and antimicrobial defense compounds. These are made from flavan-3,4-diols and -3-ols like catechins. Simpler flavonoids also tend toward 3-hydroxy types, for instance flavonols like kaempferol and quercetin, which are rarer in other plants.
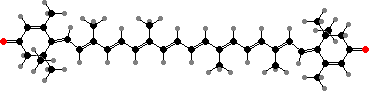
Reddish pigments here include both flavonoids and carotenoids. Rhodoxanthin in particular is found in some lycopods and horsetails, in mature leaves of most ferns, and in many conifers where it sometimes colours autumn leaves and the red seed cups of yews. It also occurs in the feathers of a few birds, though it is rare in flowering plants.
Polypodiopsida, the leptosporangiate ferns, are largely herbs from wet habitats but also include xerophytes, trees, and climbing plants. They differ from other groups in having small one-walled sporangia, normally in clusters, each formed from a single epidermal cell on the underside of the leaves. Spores develop into free gametophytes resembling non-vascular plants.
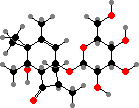
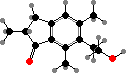
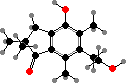
Pteridium (bracken) and some relatives have unstable glycosides like ptaquiloside. In solution these break down into pterosins, but they also readily combine with molecules like DNA, and are notable as carcinogens and livestock poisons. Various pterosins are known from many other Dennstaedtiaceae and Pteridaceae, and a few like onitin among tree ferns.
Cycadopsida, the cycads, and the following groups are all seed plants. In these the gametophytes are limited to female cells inside ovules, and male pollen, which are variously carried to the ovules and then fertilize them by means of motile gametes or a tube. The result is an embryonic sporophyte enclosed by a seed, with food reserves and a protective integument.
Cycads themselves are trees or shorter woody plants found in warmer climates. The stem is typically unbranched with a rosette of stiff, more or less pinnately compound leaves, resembling tree ferns or palms. Separate plants produce pollen cones and ovules, often pollinated by beetles, while mature seeds have a fleshy coat and are mainly dispersed by feeding mammals or birds.

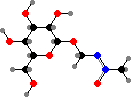
The seed coats are advertized by carotenoids, while seed kernels and leaves are protected by cycasin or related glycosides. These break down into methylazoxymethanol, which acts as an acute toxin and carcinogen through alkylation. There are a few specialist insects, though, that store the glycosides for their own defense.
Pinopsida, the conifers, are wind-pollinated trees or sometimes shrubs. The leaves are simple, either with closely branched veins or more often reduced to needles or scales. Pollen and ovules are usually formed on the same plant though in separate cones. Many grow well in cold or low-nutrient habitats and they are predominant in northern and many montane forests.
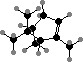
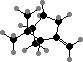
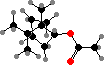
Most conifers have defensive resin consisting largely of terpenoids. The volatile components are mainly C10 hydrocarbons like α- and β-pinene enantiomers, generally with one enzyme yielding a mixture. These are particularly rich in Pinaceae, from which they are used as turpentine. The ester (–)-bornyl acetate is a main fragrance in some needle oils.
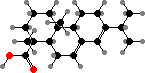
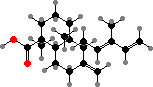
The non-volatile components are typically C20 acids, which increase the viscosity and provide a mechanical impediment to attack. Abietic acid and other tricyclic kinds are typical of Pinaceae, used as rosin. Communic acid is common in most other families; this readily forms hard polymers, preserved in class Ia and Ib ambers, the most abundant fossil resins.
Angiospermae, the flowering plants, are a diverse group ranging from minute herbs to large trees, common in nearly all terrestrial and many aquatic habitats. They differ from other seed plants in having enclosed ovules in flowers, which usually have several different types of parts, most often in whorls:
In some these are reduced or further modified. At maturity the ovaries develop into fruit of various sorts, for instance capsules which simply split open, achenes which are carried by wind, and fleshy berries and drupes which are eaten and so dispersed by animals, among others. Both flowers and fruits often attract animals with prominent colours or scents.
Most purple or blue and some red colours here, including those seen in leaves, come from anthocyanins. These are mainly glycosides of a few simple anthocyanidins, also found as minor flavonoids in other seed plants. Their form and so colour is sensitive to pH but as pigments they are typically stabilized as flavylium ions by other flavonoids, metal ions, or so on.
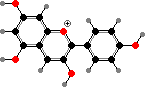
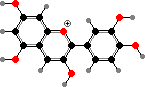
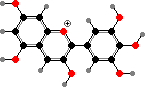
The main anthocyanidins are pelargonidin, cyanidin, and delphinidin as well as their methyl ethers. The increasing number of hydroxy groups shifts the colour to red and purple, and other interactions further to blue. Flavonols and flavones are common co-pigments, as well as forming ultraviolet markings in petals visible to insects, and giving body to white or cream flowers.
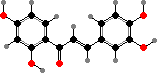
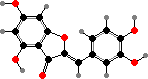
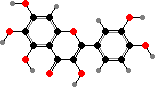
Some yellow colours in petals also come from flavonoids. The most important are chalcones like tetrahydroxychalcone 2′-glucoside and butein glucosides, which are more stable than tetrahydroxychalcone itself, and in some families brighter aurones like aureusidin. Less pale flavonols like quercetagin also contribute to colour in a few flowers.
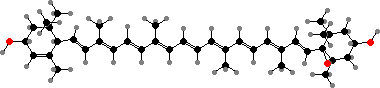
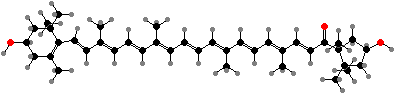
In most other flowers and fruits the main pigments are carotenoids. Yellow usually comes from fatty acid esters of lutein, other types from chloroplasts, and their epoxy derivatives. Orange or red are often from large amounts of β-carotene or lycopene, or sometimes from keto-carotenoids, notably capsanthin found in orange lilies, red peppers, and so on.
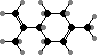
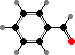
Floral scents are usually dominated by terpenoids and benzenoids. Some of the most prominent terpenoids are β-ocimene, limonenes, and linalools. These are especially associated with pollination by bees, and the last by hawk moths, while benzenoids like benzaldehyde are more associated with other insects like settling moths and butterflies.
Different angiosperms have a tremendous variety of defensive compounds, and many kinds of glycosides, alkaloids, terpenoids, and so on are connected with the diversification of different lineages. I have started on a separate angiosperm page to review them in more detail, which so far covers the following orders: